Typha angustifolia
|
Typha angustifolia L.
 (redirected from: Typha angustifolia var. calumetensis Peattie) (redirected from: Typha angustifolia var. calumetensis Peattie) |
|
|
Family: Typhaceae
Narrow-Leaf Cat-Tail, more...narrowleaf cattail, dwarf cat-tail
[Typha angustifolia var. calumetensis Peattie, moreTypha angustifolia var. elongata (Dudley) Wieg.]  Morton Arboretum |
Erect shoots 150--300 cm, not glaucous; flowering shoots 5--12 mm thick in middle; stems 2--3 mm thick near inflorescence. Leaves: sheath sides membranous, margin broadly clear, summit with membranous auricles which often disintegrate late in season; mucilage glands at sheath-blade transition brown, absent from blade and usually from sheath center near summit; widest blades on shoot 4--12 mm wide when fresh, 3--8 mm when dry; distal blade usually markedly exceeding inflorescence. Inflorescences: staminate spikes separated from pistillate by 1--8(--12) cm of naked axis, ca. as long as pistillate, 1 cm thick in anthesis; staminate scales variable in same spike, straw-colored to medium brown, filiform, simple to bifid or sometimes cuneate and irregularly branched, to 6 0.1 mm; pistillate spikes in flower when fresh dark brown with whitish stigmas (drying brown), later medium brown, in fruit when fresh as stigmas wear off often greenish due to green carpodia, (4--)6--20 cm 5--6 mm in flower, 13--22 mm in fruit; compound pedicels in fruit peg-like, 0.5--0.7 mm; pistillate bracteole blades forming spike surface before flowering, later exceeded by stigmas and about equaling or slightly exceeded by pistil hairs, very dark to medium brown, much darker than (or sometimes as dark as) stigmas, irregularly spatulate, 0.6 0.1--0.2 mm, wider than or about as wide as stigmas, apex rounded (to acute). Staminate flowers 4--6 mm; anthers 1.5--2 mm, thecae yellow, apex dark brown; pollen in monads or some in irregular clusters. Pistillate flowers 2 mm in flower, 5--7 mm in fruit; pistil-hair tips medium brown, distinctly swollen at 10--20X; stigmas sometimes deciduous in fruit, in flower erect, elongating, bending to form surface mat, white in flower, drying brownish, later medium brown, narrowly linear-lanceolate, 0.6--1.4 0.1 mm; carpodia slightly exceeded by and visible among pistil hairs at mature spike surface, green when young and fresh, straw-colored with orange-brown spots when dry, apex nearly truncate. 2n = 30. Flowering late spring--summer. Often somewhat brackish or subsaline water or wet soil; 0--1900 m; B.C., Man., N.B., N.S., Ont., P.E.I., Que., Sask.; Ark., Calif., Colo., Conn., Del., Ill., Ind., Iowa, Kans., Ky., Maine, Md., Mass., Mich., Minn., Mo., [Mont.], Nebr., Nev., N.H., N.J., N.Mex., N.Y., N.C., N.Dak., Ohio, Okla., Oreg., Pa., R.I., S.C., S.Dak., Tenn., Vt., Va., Wash., W. Va., Wis., Wyo.; Eurasia. Prior to N. Hotchkiss and H. L. Dozier (1949), Typha domingensis was generally included within T. angustifolia in North America. Because of many misidentified specimens, range expansion in recent years, and undercollecting, the distribution on the margins of the main range is somewhat uncertain. Many literature reports are based on misidentified specimens. Some workers suggested T. angustifolia was early introduced from Europe into Atlantic Coastal North America and migrated westward (R. L. Stuckey and D. P. Salamon 1987). In recent decades it has expanded its range in many regions and become much more abundant, especially in roadside ditches and other highly disturbed habitats. For example, although it was known only from one Wisconsin station in 1929 (N. C. Fassett 1930) and was very local in Iowa in 1939 (A. Hayden 1939), it is now common and widespread in both states. As it often out-competes many native marsh species to produce very dense, pure stands, and hybridizes with T. latifolia to form the probably even more competitive T. glauca, T. angustifolia and T. glauca should perhaps be classified as noxious weeds in parts of North America. Beyond the main range of T. angustifolia, there are specimens of T. glauca from north-central Montana (Phillips County.), west-central Manitoba (La Pas), and Anticosti Island, Quebec. There are m Many erroneous reports have come from outside of Europe and North America. For hybrids see also genus and key.
Perennial colony-forming herb 1 - 3 m tall Stem: erect, unbranched, 2 - 3 mm thick near the inflorescence. Leaves: sheathed at the base, long and narrow with the uppermost blade usually extending past the inflorescence, 4 - 12 mm wide when fresh and 3 - 8 mm wide when dry, convex below the middle, loosely twisted. The intersection of the sheath and blade is membranous with non-persistent ear-shaped appendages (auricles). Inflorescence: a dense cylindric terminal spike with the male part separated above the female part by 1 - 12 cm, each part about 6 - 20 cm long. The male part is 1 cm thick with straw-colored to medium brown scales that are to 6 mm long, to 0.1 mm wide, and very slender and unbranched or two-parted to wedge-shaped and branched. Female part dark brown with whitish stigmas that dry medium brown, 6 - 20 cm long, 5 - 6 mm wide, becoming 1 - 2.2 cm wide in fruit. Fruit: a tiny dry follicle borne on a slender stalk, the stalk having many long hairs with slightly widened brown tips, each follicle single-seeded. Male flowers: 4 - 6 mm long, with 1.5 - 2 mm long anthers that are yellow with dark brown tips and solitary pollen. Female flowers: subtended by dark brown spoon-shaped bracts with a rounded tip, 2 mm long, with medium brown and enlarged (visible at 10x - 20x magnification) pistil-hair tips, and white lance-linear stigmas drying to brownish. Similar species: The three most common species of Typha in the Chicago area are incredibly similar in gross morphology and one must look at the flowers and fruit (and often pollen) under magnification, and even then the identity may be inconclusive. Recently, DNA technology has become the best tool for distinguishing the taxa. While somewhat variable, the following characters may be used to separate the species, but not in all cases. Typha domingensis has light brown fruiting spikes and pointed female flower bract tips. Typha latifolia has inflorescences with the male and female parts usually touching each other, pollen grains in clusters of four, female flowers with spoon- to egg-shaped stigmas but without bracts, and more or less flattened leaves. Typha x glauca is a hybrid of T. angustifolia and T. latifolia and can often look very much like either or both parents. Certain hybrid crosses produce individuals with attributes intermediate to the parents. It is unreliable, but in the hybrid the female part of the inflorescence is often longer than 16 cm. Flowering: mid June to mid July Habitat and ecology: This species was introduced to the Chicago Region from further east and moved westward over the course of the last century. It is very common along expressways and common in marshes. It tolerates alkaline or salty soils much better than the native T. latifolia and has out-competed that species throughout the area (likely due to earlier pollen maturity). The two species hybridize to form the very aggressive and monoculture-forming Typha x glauca. Occurence in the Chicago region: non-native Etymology: Typha comes from the word typhe, the old Greek name for cat-tail. Angustifolia means narrow-leaved. Author: The Morton Arboretum Stems 1-1.5 mm; lvs 5-11 mm wide, auriculate at the juncture of sheath and blade; pistillate and staminate portions of the spike separated by (1-)2-12 cm, the pistillate portion deep brown, 10-20 cm, 1-2 cm thick at maturity; compound pedicels short and stout; pistillate fls each accompanied by a hair-like bracteole with an expanded, flat, spatulate tip; stigma linear; fr 5-8 mm, subtended by copious hairs with a slightly expanded brown tip, the achene usually distinctly above the middle; sterile pistillate fls about as long as the fertile ones, dilated into a cuneate truncate tip; staminate bracteoles brown and scale-like; pollen in monads; 2n=30. Marshes, more tolerant of salt and alkali than no. 1 [Typha latifolia L.]; nearly cosmop., and throughout our range, the common sp. along the coast, now becoming common inland as well. Gleason, Henry A. & Cronquist, Arthur J. 1991. Manual of vascular plants of northeastern United States and adjacent Canada. lxxv + 910 pp. ©The New York Botanical Garden. All rights reserved. Used by permission. From Flora of Indiana (1940) by Charles C. Deam This species is usually found on the borders of larger bodies of water than the preceding species [Typha latifolia], but it seems to adapt itself to nearly the same habitats. Near my home is a small gravel pit that has not been in use for about 10 years, and it is now filled with both species of cattails, this species occupying about a fourth of the space. It is to be noted that the pistillate part of the spike sometimes divides. I have one specimen with a 5-parted spike. I also have a specimen of the preceding species that has a 3-parted spike. This species, as well as the preceding one, is variable, and several varieties have been named. A giant form of this species is found on the east side of Tippecanoe Lake in the southern part of Noble County. Peattie's var. calumetensis seems to me to be an ecological form. Its diminutive size I attribute to the pollution of the Grand Calumet River near where it is found. In the summer when the soil along the bank is exposed it is slimy and reddish. …… Indiana Coefficient of Conservatism: C = null, non-native Wetland Indicator Status: OBL |
 Paul Rothrock  Paul Rothrock  Paul Rothrock   Biopix  Biopix  Tara Spears 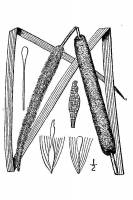 Britton, N.L., and A. Brown    Bill Harms                                                                                  |
|